Биоразнообразие / Оценка процессов биоинвазии и интродукции чужеродных видов в морских экосистемах
Перечень чужеродных видов
Бентосные организмы
Усоногие раки (Cirripedia)
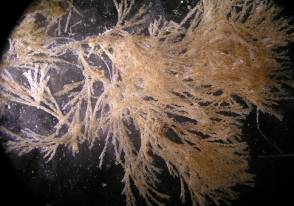
Amphibalanus improvisus. Этот вид был зарегистрирован впервые в обрастаниях гидротехнических сооружений (ГTC) зал. Петра Великого в 1969 (Зевина, Горин, 1971). Звягинцев (2003; 2005) нашел этот вид в обрастаниях всех судов каботажного и портового плавания, обследованных в заливе в период навигации - в конце июля, то есть, в начале периода, когда личинки A. impovisus начинают осаждаться (Рис. 3). В обрастаниях ГTC залива, включая Амурский зал и бухту Золотой Рог, A. improvisus был зарегистрирован почти на всех исследованных объектах как характерный вид сообществ обрастания ГTC.
Рисунок 3 – Сообщество Balanus improvisus
в судовых обрастаниях [7]
Большая экологическая пластичность и способность противостоять почти полному распреснению морской воды в бухтах и эстуариях позволили этому виду занять свободную экологическую нишу и основаться в Амурском заливе (Звягинцев, 2003). Анализ мест поселений этого вида показал, что в зал. Петра Великого развились локальные популяции A. improvisus, способные к воспроизводству и заселению не только искусственных субстратов, но также уже стали компонентом бентосных сообществ (Овсянникова, 2008; Рис. 4).
Рисунок 4 – Скопления A. Improvisus на фрагментах
древесины (А) и раковинах моллюсков (В)
(Овсянникова, 2008)
Balanus amphitrite является широко распределенным сублиторальным видом тропической и субтропической областей. Согласно Зевиной и Горину, (1975) этот вид встречается в обрастаниях буев в зал. Находка, Стрелок и Амурский только в теплые годы. В зал. Петра Великого Звягинцев (2003) обнаружил B. amphitrite в обрастаниях 46 % обследованных действующих судов за исключением плавсредств, которые провели не менее чем 20 % времени в бухте Золотой Рог, где этот вид был зарегистрирован на всех обследованных объектах. В сообществах бентоса он не встречается, однако в обрастаниях судов прибрежного и портового плавания в заливе. B. amphitrite встречаются ежегодно, а не только в теплые годы. При этом отдельные экземпляры оказались способными переносить зимние условия, однако на большинстве обследованных судов, этот вид не был зарегистрирован после зимней эксплуатации. Звягинцев (2003) не обнаружил B. amphitrite на судах портового плавания, оперирующих в портах северо-западной части Японского моря, расположенных к северу от Мыса Поворотный. Однако, этот вид обычен для обрастаний на судах прибрежного плавания, оперирующих на маршруте Владивосток – м. Сосунова (северное Приморье). Показано, что размножение и оседание личинок этого вида имеют место в период с августа по октябрь в широком диапазоне температуры воды - от 13 до 22.5°C. Взрослые особи, привнесенные в залив с судами дальнего плавания в благоприятный для их жизни сезон, продуцируют личинки, у которых имеется достаточно времени, чтобы осесть и вырасти, но это потомство погибает в зимний период.
Balanus eburneus является сублиторальным тропическо-субтропическим видом, очень уязвимым для низких температур. От был впервые обнаружен в обрастаниях мелководных хорошо прогреваемых бухт (Зевина, Горин, 1975). Эта вид может появляться в теплые годы, но не смог акклиматизировался в зал. Петра Великого и никогда не встречался в сообществах бентоса.
Balanus trigonus является тропическо-субтропическим видом, который широко распространен в теплых водах Японии. В 1970 он был найден на буях в зал. Находка (Зевина, Горин, 1975). Звягинцев (2003) обнаружил только несколько мертвых особей этого вида на танкере, который прибыл из Индии и работал в течение одной навигации на внешнем рейде Владивостока. Этот вид не смог акклиматизироваться в зал. Петра Великого.
Balanus glandula. Натурализовался в верхней сублиторали северо-восточного побережья Японских островов. Прогнозируется его появление в зал. Петра Великого [8].
Fistulobalanus albicostatus. Эта вид был найден в обрастаниях экспериментальных пластин, гидротехнических сооружений и судов (Звягинцев и др., 2009). Этот усоногий рак распространен в Японии, Корее, Китае и Калифорнии.
Megabalanus rosa. В зал. Петра Великого вид обнаружен только в биообрастаниях судов дальнего плавания. Распространен в водах Японии и Tайваня (Звягинцев и др., 2009).
Megabalanus tintinnabulum. Этот околотропический вид является видом сезонных мигрантов в зал. Петра Великого (Звягинцев и др., 2009).
Conchoderma auritium. Этот вид был найден в обрастаниях судов океанского плавания (Звягинцев и др., 2009).
Lepas anatifera. Этот вид населяет корпуса судов, буи, бакены, дрейфующий мусор, гидротехнические сооружения.
Разноногие раки (Amphipoda)
Corophium acherusicum. Этот вид был найден в зал. Петра Великого лишь в составе сообществ обрастания (Зевина, Горин 1975; Кадрьяшов, Звягинцев, 1981). Было высказано предположение, что он разносится при автотрансплантации видов A. improvisus и B. Eburneus, поселяясь в домиках умерших балянусов. Ареалы этих тропическо-субтропических видов сходны. Этот вид амфипод встречается в сообществах бентоса в юго-западной части зал. Петра Великого и вблизи устья р. Туманной (Будникова, 2001). Этот вид встречается на шельфе восточного Сахалина, а также в обрастаниях системы охлаждения ТЭЦ-2 г. Владивостока (Звягинцев, Будникова, 2003). Это является свидетельством натурализации C. acherusicum в зал. Петра Великого.
Равноногие раки (Isopoda)
Sphaeromatiidae. Вид был обнаружен в биообрастаниях судов, предположительно следующих из портов Австралии или Сингапура (Ростомов, 1981).
Крабы и раки-отшельники (Decapoda)
Planes marinus. Этот краб был обнаружен на дрейфующем буе в зал. Петра Великого. Вероятный источник – вторжения субтропических вод в залив в летний период (Кепель и др., 2002).
Portunus sanguinolentus. Этот краб был также найден в зал. Петра Великого на дрейфующем буе (Рис. 5). Высказано предположение, что появление представителей тропической фауны у побережья Приморья связано с увеличение количества морского мусора, переносимого морскими течениями, который служит субстратом для поселения различных морских организмов (Кепель, Царева, 2005).
Plagusia depressa tuberculata. Этот краб также был обнаружен в зал. Петра Великого на дрейфующем буе (Рис.6). Его появление здесь обусловлено теми же причинами, что и предыдущего вида.
Diogenes nitidimanus. Этот рак-отшельник впервые был обнаружен в зал. Востока (зал. Петра Великого) на илистом основании на глубине 2-3 м (Корн и др., 2007, Рис. 7). Этот вид заселяет раковины брюхоногих моллюсков. Позже, личинка рака-отшельника были найдена в балластных водах танкера, прибывшего из Китая.
Рисунок 7 - Вид-вселенец Diogenes nitidimanus
Гидроиды (Hydroidea)
Gonothyrea loveni. Этот вид гидроида, известный в Северной Атлантике, был впервые зарегистрирован и в обрастаниях судов в Татарском проливе, а позже – ГТС (Чаплыгина, 1980). Считается, что G. loveni находится здесь на первой стадии акклиматизации, так как он не зарегистрирован в сообществах бентоса и его количественные показатели невысоки [7].
Campanularia johnstoni. Этот вид широко распространен в обоих полушариях, исключая районы высоких широт. Согласно Багавеевой с соавторами (1984) этот вид уже натурализовался в донных сообществах северо-западной части Японского моря.
Laomedea flexuosa. Естественным ареалом обитания этого вида обычно являются воды Северной Атлантики и некоторых северных морей России. Обнаружен в обрастаниях судов дальнего плавания, прибрежного судоходства, а также пирсов и причалов в зал. Петра Великого и северных прибрежных районов Приморья.
Laomedea calceolifera. Естественный ареал обитания этого вида ранее охватывал умеренные воды Северной Атлантики. В течение последних двух десятилетий он обнаруживается в обрастаниях пирсов и причалов зал. Петра Великого, а также пос. Пластун (Чаплыгина, 1992). Однако о натурализации здесь этих последних двух видов говорить пока преждевременно [7].
Aequorea coerulescens. Этот вид был впервые зарегистрирован в приморских водах в 1995 (Погодин, Яковлев, 1999). Он известен в Японии, включая северное побережье о. Хоккайдо. Однако неясно, является ли он чужеродным видом, или это просто его первое открытие в недостаточно изученном районе.
Многощетинковые черви (Polychaeta)
Polydora limicola. Распространен в бентосе Берингова моря, Авачинской губы, п-ва Шикотан и Калифорнийского побережья. В Японском море этот вид встречается в обрастаниях ГТС Владивостока и Находки (Багавеева, 1981). В бентосе он не обнаружен. Вид обладает высокой экологической пластичностью, переносит значительное понижение солености и загрязнение вод нефтепродуктами. По-видимому, в северо-западной части Японского моря этот вид находится в первой стадии акклиматизации на акваториях портов.
Hydroides elegans. Это эврибионтный вид, достаточно устойчивый к колебаниям солености и сильному загрязнению (Багавеева, 1981 и др.). H. elegans был найден в в зал. Петра Великого в обрастаниях судов и экспериментальных пластин, установленных в бухте Золотой Рог (Горин, 1975). Этот вид доминирует среди обрастателей в заливе, и его биомасса в целом увеличивается, особенно по направлению к кутовой части бухты.
Pseudopotamilla occelata. В естественных условиях это вид обитает на литорали и сублиторали Аляск, Орегона, Калифорнии и Японии (Рис. 8). В северо-западной части Японского моря был впервые обнаружен в 1980 в обрастаниях гидротехнических сооружений нескольких портов и портопунктов остовов и заливов от зал. Посьета на юге – до Холмска на севере (Звягинцев, 2003). Основной источник распространения – судоходство. Его натурализация в зал. Петра Великого привела к существенным изменениям в составе бентосных сообществ и свидетельствует об угрозе серьезных экологических последствий, включая опасности колоний Р. оccelata для здоровья человека. Вместе с тем, натурализация этого вида имеет и определенное практическое значение, т. к. массовое развитие полихет положительно отражается на динамике отдельных видов рыболовства [7].
В презентации [11] упоминаются и некоторые другие виды полихет, интродуцированных в залив Петра Великого (согласно рис. 9).
Mytilus galloprovinciallis. Этот вид был обнаружен в зал. Посьета, юго-западной части зал. Петра Великого в начале 1980-х гг. на плавающих предметах и был идентифицирован как “Mytilus edulis diegensis”. Дальнейшие исследования показали, что это - M. galloprovincialis (Рис. 10), имеющий средиземноморское происхождение, скорее его гибрид с M. trossulus, и он уже распространился в нескольких районах зал. Петра Великого и севернее. (Иванова, Лутаенко, 1998).
Gomphina aequilatera. Этот вид был обнаружен как пустые раковины в пляжных наносах (Рис. 11) вблизи российско-корейской границы в 1997 (Лутаенко, Яковлев, 1999; Лутаенко, 2007). Обычно он распространен в субтропических водах и ранее никогда не регистрировался в бухтах зал. Петра Великого.
Haliotis discus. Этот вид (Рис. 12) – обычный обитатель субтропических и тропических вод. Ранее он не регистрировался в зал. Петра Великого несмотря на долгую историю зоологических и гидробиологических исследований на его акватории. В 2002 г. он был впервые обнаружен на подводных скалах вблизи русско-корейской на глубине 13 м. (Раков, Архипов, 2004), а позже – вблизи о. Монерон у южного Сахалина.
Aplysia parvula. Этот вид “морского зайца” (Рис. 13) впервые был найден у о. Фуругельма, на юге зал. Петра Великого в 2005 (Чернышев и др., 2006) Его появление можно объяснить перемещением личинок с водами теплых течений.
Рисунок 13 - Вид-вселенец Aplysia parvula (Чернышев и др., 2006)
Cellana toreuma. Этот субтропический вид был обнаружен в единственном экземпляре в пляжных наносах о. Фуругельма (Чернышев, Чернова, 2003). Его также можно рассматривать как возможную инвазию.
Alderia modesta. Моллюск был найден в поселениях водорослей Vaucheria в одной из лагун Амурского залива. В конце мая - начале июня плотность поселений A. modesta достигала 2000 экземпляров/м² (Чернышев, Чабан, 2005). Его также можно рассматривать как возможную инвазию.
Мшанки (
yozoa)
Bugula californica. Этот вид (Рис. 14) из тихоокеанских североамериканских вод обнаружен в обрастаниях судов прибрежного плавания в северо-западной части Японского моря. B. californica найден на 41 % судов, осмотренных в зал. Петра Великого. В бухте Золотой Рог он выявлен на 83 % судов в сообществе H. elegans. В сообществах мидий и устриц вид B. californica обнаружен на 69 и 50 % исследованных судов, соответственно. Этот вид не был зарегистрирован на судах, работающих к северу от м. Поворотный, и пока находится в первой стадии акклиматизации (Звягинцев, 2003).
Рисунок 14 - Вид-вселенец Bugula californica [11]
Conopeum seurati. Этот вид распространен на атлантическом побережье Европы, от Британских островов до Средиземного и Черного морей, откуда расселился в другие районы. Обнаружен в составе донных сообществ заливов Посьета и Восток, а также в б. Золотой Рог и на буе у м. Улисс (Зевина, Горин 1975). C. seurati найден и в обрастаниях 65% судов, не выходящих за м. Поворотный. Максимальная биомасса этого вида (12 гр/м²) была зарегистрирована на судах, находящихся или работающих в б. Золотой Рог (Звягинцев, 2003). Вселение этого вида может служить классическим примером интродукции с последующими акклиматизацией и натурализацией.
Schizoporella unicornis. Этот вид входит в состав сообществ обрастания причалов зал. Посьета, широко распространен в бентосе верхней сублиторали заливов Посьета и Восток (Зевина и др., 1975). Обнаружен на 25 % судов, обследованных в зал. Петра Великого при незначительной биомассе и частоте встречаемости в пробах 6 %. Севернее м. Поворотный отмечен только в пос. Преображение (Звягинцев, 2003).
Bowerbankia gracilis. Багавеева с соавторами (1984) полагают, что этот вид был интродуцирован автотрансплантацией и уже натурализовался в зал Петра Великого. Возможно, что он является аборигеном северо-западной части Японского моря, который плохо переносит условия существования в портовых водах [7].
Асцидии (Tunicata)
Molgula manhattensis. Естественный ареал этого вида находится в водах прибрежной зоны Северной Америки, откуда он быстро распространяется по всему свету. Этот вид быстро приспосабливается к водам пониженной солености и может встречаться в закрытых водоемах с сильно опресненной и загрязненной взвесью водой. Впервые был обнаружен в сообществах обрастания пластин в б. Золотой Рог и б. Рында в 1999 г. (Рис. 15). При дальнейшем распространении вида могут возникнуть серьезные проблемы при эксплуатации гидротехнических сооружений и подвесной марикультуры [8].
Ciona savignyi. Естественными районами обитания вида (Рис. 16) являются умеренные широты Тихого океана. В зал. Петра Великого впервые был обнаружен в 2004 г. в зал. Восток, где ранее не встречался, в начале лета на глубине 2-4 м на остатках рыболовных сетей, а также в сообществе мидий на корпусе плавучего дока в б. Гайдамак. Количественные показатели распространения вида на новом местообитании достаточно высоки. При дальнейшей интродукции и акклиматизации вида в зал. Петра Великого может возникнуть проблема зарастания подвесных садков в хозяйствах марикультуры [7].
Botrylloides diegense. Этот вид был обнаружен в зал. Петра Великого в 1969 г., куда от был интродуцирован из района южной Калифорнии (Беньяминсон, 1981).
Botryllus tuberatus. Этот вид был обнаружен в донных сообществах зал. Посьета на глубинах 1,5 – 11 м. (Беньяминсон, 1981). Распространен в прибрежной зоне морей западной экваториальной зоны Тихого океана.
Polyandrocarpa zorritensis. Естественный ареал этого вида находится в водах прибрежной зоны Северной Америки, откуда он распространился до Японских островов. Прогнозируется его появление в сообществах обрастания зал. Петра Великого [8].
Планктонные организмы и грибы
В ходе исследований балластных вод двух судов на Российско-Японской и Российско-Китайской линиях [12] было обнаружено 45 видов микроводорослей, 22 вида зоопланктона, 11 видов меропланктона, 10 групп мейофауны, идентифицировано 24 вида из микроскопических мицелиальных грибов, в культуры из двух проб было выделено 28 и 12 морфологически отличающихся штамма бактерий – всего не мене 152 видов. Так балластных танках сухогруза «Timber Star» обнаружены потенциально токсичные виды Pseudo-nitzschia pungens, Chaetoceros sp., Asterionellopsis glacialis, Sceletonema costatum, Dinophysis acuminata, Prorocentrum cordatum, Dino
yon balticum, Euglena sp., Eutreptiella eupharyngea – в основном космополиты, массовое развитие которых в заливе Петра Великого отмечают в летне-осеннее время (Стоник, Орлова, 1998).
Голопланктон балластных вод состоял из субтропических и бореальных видов, относящихся к Copepoda, Cladocera, Mysidacea, Chaetognatha и Hydrozoa. Фауна балластных вод была представлена неритическими (71%) и океаническими видами (29%). Веслоногие ракообразные Copepoda были представлены 9 планктонными видами, а также обитателями придонного слоя (отр. Harpacticoida). Из ветвистоусых ракообразных Cladocera в заметных количествах встречались как эврибионтные Evadne nordmanni, холодноводные Podon leuckarti, Pseudocalanus newmani, Centropages abdominalis, Acartia hudsonica, так и нетипичные редкие тепловодные виды Pseudevadne tergestina и Pseudocalanus inopinus. P. inopinus – эндемик тропической области Индо-Тихоокеанского региона. В заливе Петра Великого эта «азиатская копепода» встречается единично, в основном в его южной части, и является индикатором поступления теплых вод с юга. При натурализации в эстуарные зоны он способен вытеснять другие виды, изменяя структуру планктонного сообщества (Bollens, Murphy, Ritz, Hewitt, 2002).
В мае и, как правило, в конце июля (при прогреве вод выше 17 °С) в заливе отмечены виды тропического происхождения Oithona plumifera и O.
evicornis (Касьян, Чавтур, 2006).
В балластных водах обнаружено и идентифицировано 24 вида морских грибов из 10 родов микроскопических мицелиальных грибов. Они представлены анаморфными грибами (Anamorphic Fungi) – 22 вида (91.7%) и зигомицетами (Zygomycota) – 2 вида (8.3%). Доминировали представители родов Aspergillus – 7 видов (29.2%), Penicillium – 6 видов (25%), Acremonium – 3 вида (12.5%), Cladosporium – 2 вида (8.3%). По 1 виду (≈ 4.2%) обнаружено в родах Mucor, Rhizopus, Alternaria, Fusarium, Geomyces и Trichoderma.
тивную стадию и спороношения [12].
Мицелиальные грибы– представители родов Aspergillus (A. niger, A. flavus, A. ochraceus, A. versicolor), Penicillium, Cladosporium (C. sphaerospermum), Alternaria (A. alternata), Trichoderma, Mucor (M. racemosum), Rhizopus (Rh. nigricans), обнаруженные в балластных
водах, относятся к группе условно-патогенных грибов и известны как возбудители глубоких и оппортунистических микозов как у человека и наземных животных, так и у морских беспозвоночных и рыб (Sallenave-Namont et al., 2000; Sindermann, Lightner, 1988).
Виды рода Fusarium вызывают инфекции молоди культивируемых ракообразных (Sindermann, Lightner, 1988). Выявленные мицелиальные грибы являются продуцентами гемолитических метаболитов и антибиотиков, что обуславливает их патогенные свойства и конкурентоспособность с другими микроорганизмами (Pivkin, 2000).
Нектонные и нектобентосные организмы
Медузы (Scyphozoa)
Rhopilema esculentum. Этот вид медузы распространен в водах тропических и субтропических морей – Южно-Китайском, Желтом и Восточно-Китайском. Его проникновение в зал. Петра Великого связывается с интенсификацией потоков теплых Цусимского и Восточно-Корейского течений (Бородин и др., 2003).
Рыбы (Pisces)
Комплексный анализ ихтиофауны зал. Петра Великого за вековой период выявил 310 видов рыб, относящихся к 24 отрядам, 83 семействам, 204 родам. Общее число рыб, отмеченных в заливе по десятилетним интервалам наблюдений, колебалось от 232-233 видов в периоды потеплений до 198-205 видов в периоды похолоданий [13].
Южные мигранты – 106 видов – играют значительную роль в видовом разнообразии рыб залива (Рис. 17). Больше всего тепловодных видов рыб было зарегистрировано в первой половине ХХ века, во время первой волны потепления – до 55 видов в десятилетнем интервале наблюдений. В 1990-е годы, когда наблюдалась вторая, более сильная волна потепления, число южан было также значительным – до 53 видов.
Количество и состав рыб-мигрантов в водах зал. Петра Великого определяются рядом факторов, из которых на первом месте пока остается климатический фактор при все возрастающей роли антропогенного. Биотический фактор также в определенные периоды усиливает миграционную активность рыб, особенно хищников-преследователей.
Изменения в ихтиофауне залива, особенно в группе южных мигрантов, затронули не только количественный состав, но и структуру ихтиофауны. Уменьшилось число видов рыб отрядов Lamniformes,Rajiformes, Tetraodontiformes в результате как прямого перелова, так и разрушения сложившихся условий существования и воспроизводства. В первой половине ХХ века больше отмечалось хищных рыб крупных размеров. В настоящее время увеличение видового разнообразия идет в основном за счет мелких прибрежных видов из отрядов Perciformes и Scorpaeniformes.
Северные мигранты в заливе представлены 28 видами, большее число их было зарегистрировано в водах в начале ХХ в., как результат предшествовавшего длительного похолодания. К концу столетия, в связи с потеплением, их число сократилось до 2-4 видов в десятилетних интервалах наблюдений.
Разнообразие ихтиофауны зал. Петра Великого в условиях потепления сохранится на достаточно высоком уровне. В ближайшие годы, если тренд потепления продлится, можно ожидать увеличения числа южных мигрантов, в том числе появления новых для вод залива и России видов.
Чрезмерный промысел привел к снижению численности сельди, камбал, наваги и других рыб-резидентов залива, в их числе и анадромных видов – горбуши, кеты, симы, корюшки-зубатки.
В результате хозяйственной деятельности в реки, впадающие в зал. Петра Великого, интродуцировано 12 (13) видов рыб. Влияние и роль их на видовой состав «местных» рыб еще не до конца выяснено (в соответствии с [13]).
Более подробные количественные сведения по видам-мигрантам приводятся в разделе информационной системы «Оценка динамики изменений ихтиофауны зал. Петра Великого».
Рисунок 17 – Отдельные представители видов рыб – южных мигрантов в зал. Петра Великого [16]
Морские рептилии (Chelonia and Serpentes)
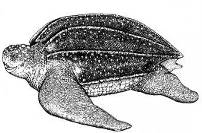
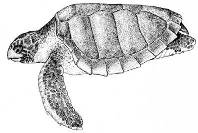
Морские рептилии (черепахи и морские змеи) обнаруживались в российских водах Японского моря с конца 19-го столетия (1873 г.) и в последующие годы. Это желтобрюхая морская змея Pelamis platura (1873, 2007) (Рис. 18), китайская морская змея Pseodolaticauda semifasciata (1978 г.), кожистая черепаха Dermochelys coriacea (1936 г.) (Рис. 19) и черепаха вида Caretta caretta (1940 г.) (Рис. 20) (Харин, 2008). Основная причина миграции этих теплолюбивых видов в северном направлении – интенсификация теплых течений и прогрев прибрежных вод зал. Петра Великого.
Ожидается, что с учетом тенденций глобальных климатических изменений, количество подобных и других чужеродных видов в перспективе здесь может значительно увеличиться.
 Amphibalanus improvisus. Этот вид был зарегистрирован впервые в обрастаниях гидротехнических сооружений (ГTC) зал. Петра Великого в 1969 (Зевина, Горин, 1971). Звягинцев (2003; 2005) нашел этот вид в обрастаниях всех судов каботажного и портового плавания, обследованных в заливе в период навигации - в конце июля, то есть, в начале периода, когда личинки A. impovisus начинают осаждаться (Рис. 3). В обрастаниях ГTC залива, включая Амурский зал и бухту Золотой Рог, A. improvisus был зарегистрирован почти на всех исследованных объектах как характерный вид сообществ обрастания ГTC.
Amphibalanus improvisus. Этот вид был зарегистрирован впервые в обрастаниях гидротехнических сооружений (ГTC) зал. Петра Великого в 1969 (Зевина, Горин, 1971). Звягинцев (2003; 2005) нашел этот вид в обрастаниях всех судов каботажного и портового плавания, обследованных в заливе в период навигации - в конце июля, то есть, в начале периода, когда личинки A. impovisus начинают осаждаться (Рис. 3). В обрастаниях ГTC залива, включая Амурский зал и бухту Золотой Рог, A. improvisus был зарегистрирован почти на всех исследованных объектах как характерный вид сообществ обрастания ГTC.